動物細胞構造
動物細胞は、原形質膜によって囲まれ、膜結合核および細胞小器官 植物や真菌の真核細胞とは異なり、動物細胞は細胞壁を持っていません。 この特徴は、動物界の王国を生み出した単細胞生物によって遠い過去に失われました。, ほとんどの細胞は、動物および植物両方、1そして100マイクロメートルの間で、及びこうして顕微鏡の援助とだけ目に見えます。
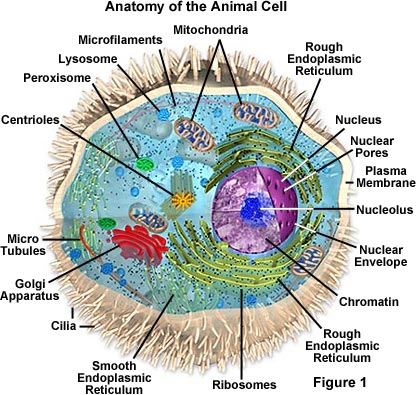
硬い細胞壁の欠如は、動物が細胞型、組織、および器官のより多くの多様性を発達させることを可能にした。 神経や筋肉を形成する特殊な細胞—植物が進化することができない組織—は、これらの生物に移動性を与えました。, 特殊な筋肉組織の使用によって動き回る能力は、動物界の特徴ですが、いくつかの動物、主にスポンジは、分化した組織を持っていません。 特に、原生動物は移動するが、それは繊毛、鞭毛、および偽足を使用して、実際には、非筋肉的手段を介してのみである。
ほとんどの動物組織は、コラーゲンとして知られているタンパク質の三重らせんによって細胞外マトリックス内で一緒に結合されているため、動物界は真核生物の間でユニークです。 植物細胞および真菌細胞は、ペクチンなどの他の分子によって組織または凝集中で一緒に結合される。, 他の生物がこのようにコラーゲンを利用していないという事実は、すべての動物が共通の単細胞の祖先から生じたという兆候の一つです。 動物細胞間のコラーゲン含有細胞外マトリックスが石灰化すると、骨、殻、spicules、および他の硬化構造が形成される。
動物は生物の大規模かつ非常に多様なグループです。 地球上の種の約四分の三を構成し、彼らはアリ、クジラ、ゾウ、そしてもちろん、人間にサンゴやクラゲから色域を実行します。, モバイルであることは、彼らの環境を感知し、応答することができる動物、供給、防衛、および再生の多くの異なるモードを採用する柔軟性を与えています。 しかし、植物とは異なり、動物は自分の食物を製造することができないため、常に直接的または間接的に植物の生命に依存しています。
ほとんどの動物細胞は二倍体であり、それらの染色体は相同対に存在することを意味する。 しかしながら、異なる染色体の倍数性も時折起こることが知られている。 動物細胞の増殖は様々な方法で起こる。, 有性生殖の例では、減数分裂の細胞プロセスが最初に必要であり、一倍体の娘細胞または配偶子を産生することができる。 二つの一倍体細胞は、その後、その細胞が分裂し、乗算として新しい生物に発展二倍体接合体を形成するために融合します。
動物の最も初期の化石証拠はヴェンディアン時代(650万年から544万年前)にさかのぼり、浅い水の堆積物に柔らかい体の痕跡を残した腔内型の生き物である。, 最初の大量絶滅はその期間を終えましたが、それに続くカンブリア紀の間に、新しい形態の爆発が今日知られている主要なグループ、または門のほとんどを生み出した進化の放射を始めました。 脊椎動物(背骨を持つ動物)は、初期のオルドビス紀(505-438万年前)まで発生していることが知られていません。
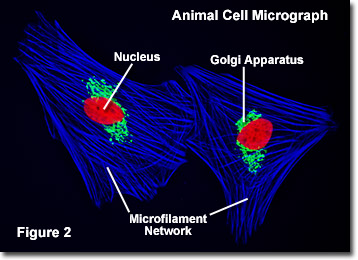
細胞は、1665年に英国の科学者ロバート-フックによって発見されました。, 実際には、フックは、彼が小さな、裸の部屋や僧侶の細胞のようなコルクの微視的な構造を記述したときに、生物学的文脈で、用語”細胞”を造語しました。 図2に示すのは、蛍光プローブで標識され、その内部構造を明らかにするために顕微鏡で撮影された一対の線維芽細胞シカ皮膚細胞である。 核は赤色プローブで染色され,ゴルジ装置とミクロフィラメントアクチンネットワークはそれぞれ緑色と青色で染色された。, 顕微鏡は、細胞生物学の分野における基本的なツールであり、培養中の生きた細胞を観察するためにしばしば使用される。 以下のリンクを使用して、動物細胞に含まれるさまざまな成分に関するより詳細な情報を入手してください。
-
セントリオール-セントリオールは、微小管の九つの束からなる自己複製オルガネラであり、動物細胞にのみ見出される。 彼らは細胞分裂を組織するのに役立つように見えますが、プロセスに不可欠ではありません。,
-
繊毛および鞭毛-単細胞真核生物にとって、繊毛および鞭毛は、個々の生物の移動に不可欠である。 多細胞生物では、繊毛は、不動の細胞を過ぎて流体または物質を移動させるだけでなく、細胞または細胞群を移動させる機能を果たす。
-
小胞体-小胞体は、細胞の内部および外部で使用するための化合物を製造、処理、および輸送する嚢のネットワークである。 これは、核と細胞質との間のパイプラインを提供し、二重層核エンベロープに接続されています。,
-
エンドソームとエンドサイトーシス-エンドソームは膜結合小胞であり、エンドサイトーシスと呼ばれる複雑なプロセスファミリーを介して形成され、ほぼすべての動物細胞の細胞質に見られる。 発現のメカニズムは、基本的にエンドサイトーシスが、逆の発生時の逐次開口放出や細胞分泌である。 これは、細胞の原形質膜の陥入(内側に折り畳まれる)を伴い、細胞外液を介して拡散する高分子または他の物質を取り囲む。,
-
ゴルジ装置-ゴルジ装置は、細胞の化学製品の流通および出荷部門です。 それは小胞体で造られる蛋白質および脂肪を変更し、細胞の外側に輸出のためにそれらを準備します。
-
中間フィラメント-中間フィラメントは、細胞骨格の構造的および機能的要素の両方として重要な役割を果たす繊維タンパク質の非常に広い に至るまでのサイズから8-12ナノメートル(中間径フィラメントとして機能し、張力負の要素を維持するための細胞形状を高い生産性が可能となります。,
-
リソソーム-これらのマイクロボディの主な機能は消化です。 リソソームは、細胞の外部からの細胞廃棄物や破片を単純な化合物に分解し、新しい細胞構築材料として細胞質に移します。
-
ミクロフィラメント-ミクロフィラメントは、アクチンと呼ばれる球状タンパク質からなる固体棒です。 これらのフィラメントは機能で主に構造で、細胞骨格の重要な部品です。,
-
微小管-これらの真っ直ぐな中空の円柱は、すべての真核細胞の細胞質全体に見られ(原核生物にはそれらがない)、輸送から構造支持までの様々な機能
-
ミトコンドリア-ミトコンドリアは、すべての真核細胞の細胞質に見出される楕円形の細胞小器官である。 動物細胞では、それらは主発電機であり、酸素と栄養素をエネルギーに変換します。,
-
核-核は、細胞の情報処理および管理センターとして機能する高度に特殊化されたオルガネラである。 このオルガネラは、細胞の遺伝物質、またはDNAを格納し、それが成長、中間代謝、タンパク質合成、および再生(細胞分裂)を含む細胞の活動を調整する:二つの大きな機能を持っています。
-
ペルオキシソーム-マイクロボディーは、細胞質に見られ、ほぼ球形で、単一の膜によって結合された多様なオルガネラ群である。, マイクロボディにはいくつかの種類がありますが、ペルオキシソームが最も一般的です。
-
原形質膜-すべての生きている細胞は、その内容物を囲む原形質膜を有する。 原核生物では、膜は堅い細胞壁によって囲まれる保護の内部の層です。 真核動物細胞は、それらの内容物を含み、保護する膜のみを有する。 これらの膜はまた、細胞の中および外の分子の通過を調節する。
-
リボソーム-すべての生きている細胞は、リボソーム、約60%のRNAと40%のタンパク質からなる小さな細胞小器官を含んでいます。, 真核生物では、リボソームはRNAの四本の鎖から成っています。 原核生物では、それらはRNAの三本鎖からなる。
さらに、光学および電子顕微鏡では、科学者は動物細胞の謎を調べるために他の多くの技術を使用することができます。 細胞は化学的方法によって分解することができ、そしてそれらの個々の細胞小器官および高分子は研究のために単離される。 細胞分画のプロセスは、科学者がその組成および機能の調査のために、例えばミトコンドリアなどの特定の成分を大量に調製することを可能にする。, このアプローチを使用して、細胞生物学者は、細胞内の特定の場所に様々な機能を割り当てることができました。 しかし、蛍光タンパク質の時代は、生命プロセスの微妙なバランスを妨げない研究のために、高度に局在化したプローブで生きた細胞を標的とすることを可能にすることで、顕微鏡を生物学の最前線にもたらしました。
細胞構造ホームに戻る
細胞の蛍光顕微鏡に戻る
質問やコメント? 私達に電子メールを送りなさい。
©1995-2019by Michael W., デビッドソンとフロリダ州立大学。 すべての権利を保有します。 著作権者の許可なく、画像、グラフィック、ソフトウェア、スクリプト、またはアプレットを複製または使用することはできません。 このウェブサイトの使用は、あなたが所有者によって定められた法的条件のすべてに同意することを意味します。
このウェブサイトは、私たちによって維持されています
グラフィックス&ウェブプログラミングチーム
国立高磁場研究所の光学顕微鏡,
Last modification: Friday, Nov 13, 2015 at 01:18 PM
Access Count Since October 1, 2000: 6031106
Microscopes provided by:
![]()
![]()
Leave a Reply